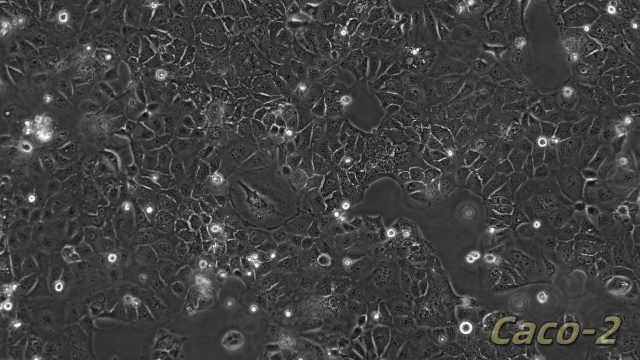
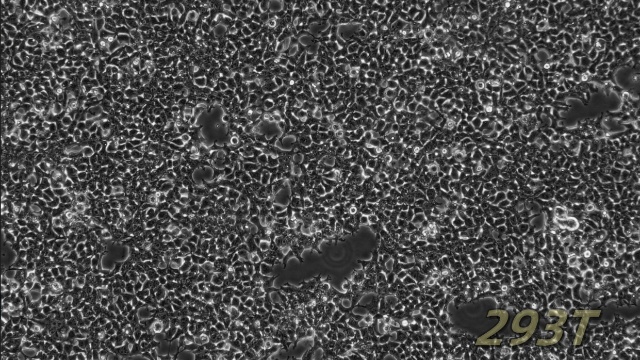
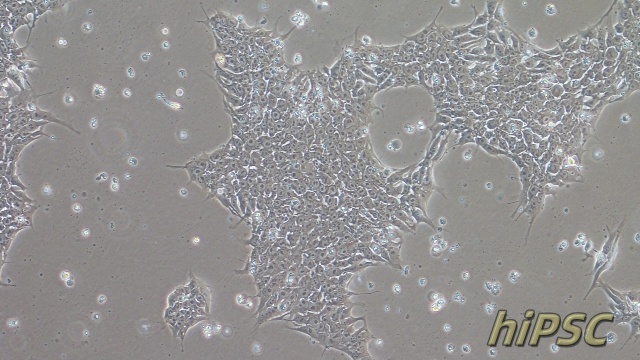
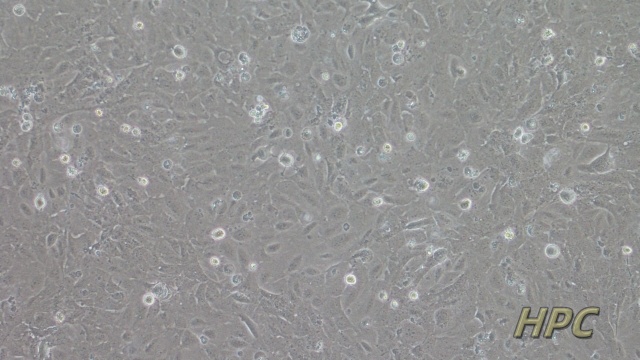
NCM460人正常结肠上皮细胞是一种来源于正常人结肠组织的细胞系,广泛应用于肠道生物学和消化系统研究领域。该细胞系保留了结肠上皮细胞的典型特性,能够表达肠道特异性标志物(如紧密连接蛋白和黏蛋白),并具备结肠上皮细胞的屏障功能和分泌功能。NCM460细胞在体外培养中表现出稳定的增殖能力和功能活性,常用于研究肠道屏障机制、营养物质吸收以及结肠上皮细胞对外界刺激的响应。由于其对人结肠上皮细胞功能的良好模拟,NCM460细胞成为研究肠道生理功能、炎症反应以及肠道相关信号通路的重要模型。此外,NCM460细胞在药物筛选、毒性测试以及肠道微生态相互作用研究中也发挥了重要作用。由于其易于培养和广泛的应用价值,NCM460人正常结肠上皮细胞为肠道生物学研究提供了重要的实验工具,为深入理解肠道功能和相关机制提供了有力支持。细胞内的细胞间连接结构维持组织完整性。SVPVERO细胞衍生株
RSC96细胞是一种来源于大鼠的雪旺细胞系,雪旺细胞是周围神经系统中的重要胶质细胞,主要负责形成髓鞘并支持神经元的正常功能。RSC96细胞在体外培养中表现出典型的雪旺细胞形态和功能特性,是研究周围神经发育、髓鞘形成及神经再生的常用模型。通过研究RSC96细胞,可以深入探讨雪旺细胞在神经损伤修复中的作用机制,例如细胞外基质相互作用、神经营养因子的分泌以及髓鞘相关蛋白的表达调控。此外,RSC96细胞还被用于研究雪旺细胞与神经元之间的相互作用,揭示其在神经信号传导和维持神经微环境中的关键功能。由于其易于培养且稳定性较高,RSC96细胞在神经生物学研究中具有重要价值,为探索周围神经系统的生理和病理机制提供了有力工具。重庆细胞有哪些细胞内的蛋白质降解系统维持蛋白质稳态。

3T3-L1小鼠胚胎成纤维细胞是一种***用于脂肪细胞分化研究的细胞系,起源于Swiss3T3小鼠胚胎。该细胞具有典型的成纤维细胞形态,贴壁生长,能够在特定诱导条件下分化为成熟的脂肪细胞,因此成为研究脂肪生成、脂质代谢和胰岛素信号通路的经典模型。在分化过程中,3T3-L1细胞经历从成纤维细胞样形态向圆形脂肪细胞样形态的转变,并积累脂滴。分化诱导通常采用含有胰岛素、**和3-异丁基-1-甲基黄嘌呤(IBMX)的培养基,***PPARγ和C/EBPα等关键转录因子,驱动脂肪生成相关基因的表达。分化后的细胞表现出典型的脂肪细胞特性,如脂质储存和***敏感性。3T3-L1细胞在代谢疾病研究中具有重要价值。例如,它们被用于研究肥胖、2型糖尿病和非酒精性脂肪肝等疾病的分子机制。通过基因编辑技术(如CRISPR-Cas9)或药物处理,科学家可以模拟疾病状态,探索新的***靶点。此外,3T3-L1细胞还被用于筛选调节脂质代谢和胰岛素敏感性的化合物,为开发代谢疾病***药物提供了重要平台。
MEF(Mouse Embryonic Fibroblasts)小鼠胚胎成纤维细胞是一种来源于小鼠胚胎的细胞系,广泛应用于发育生物学、细胞生物学和再生医学研究领域。该细胞系具有典型的成纤维细胞特性,能够分泌多种细胞外基质成分,并参与组织修复和再生过程。MEF细胞在体外培养中表现出稳定的增殖能力和多向分化潜能,常用于研究细胞增殖、分化、衰老以及细胞外基质的相互作用。由于其来源***且易于培养,MEF细胞成为研究胚胎发育、干细胞微环境以及细胞信号通路的理想模型。此外,MEF细胞在基因功能研究、药物筛选以及组织工程研究中也发挥了重要作用。由于其多功能性和广泛的应用价值,MEF小鼠胚胎成纤维细胞为发育生物学和再生医学研究提供了重要的实验工具,为深入理解细胞行为和发育机制提供了有力支持。细胞分裂是生物体生长和繁殖的基础。

HHDPC(人毛**细胞)是位于***基底部的特殊间充质细胞,在毛发生长周期中发挥重要的调控作用。这些细胞通过分泌多种生长因子和信号分子,参与***的形成、维持以及再生过程。HHDPC在体外培养中能够保持其独特的生物学特性,是研究毛发生长机制和***生物学的重要模型。通过研究HHDPC,可以深入探讨***发育和周期性生长的分子机制,例如细胞外基质的相互作用、生长因子的表达调控以及细胞间信号通路的***。此外,HHDPC还被用于研究毛**细胞与***上皮细胞之间的相互作用,揭示其在***微环境中的**作用。由于其来源明确且功能独特,HHDPC在毛发相关研究中具有重要价值,为探索毛发生理和再生机制提供了重要工具。细胞内的液泡储存水分和营养物质。CGM1人EB病毒转化的B细胞
细胞通过内吞作用摄取大分子物质。SVPVERO细胞衍生株
C6/36细胞系是从白纹伊蚊(Aedesalbopictus)胚胎中分离的连续传代细胞,广泛应用于蚊媒病毒学研究。该细胞具有典型的贴壁生长特性,在28℃无CO₂条件下可稳定增殖,适合多种虫媒病毒的培养,包括登革病毒、寨卡病毒和基孔肯雅病毒等。其突出的病毒敏感性源于蚊源细胞天然的病毒复制支持系统,为病毒-宿主互作机制研究提供了理想模型。在操作流程上,C6/36细胞常用Leibovitz'sL-15培养基培养,需定期传代维持密度在70%-90%。值得注意的是,该细胞存在缺陷型干扰颗粒积累现象,长期传代可能导致病毒产量下降,建议控制传代次数。近年研究发现,其对某些病毒株的***会引发***细胞病变效应,如细胞圆缩和脱落,这为病毒致病性研究提供了可视化指标。该细胞系还被用于探索蚊虫抗病毒免疫通路,如RNA干扰机制在限制病毒复制中的作用。通过比较不同代次细胞的转录组差异,科研人员可追踪体外培养对细胞特性的影响。因其保留部分蚊体组织特性,在媒介生物学领域具有独特价值。SVPVERO细胞衍生株